Module 2.3 Modélisation spatiale
Résumé du module
Dans ce module, nous vous présentons les concepts importants de la modélisation spatiale. Ce module comprend quatre activités : trois activités de lecture et une activité d’exploration de modèle avec le logiciel NetLogo.
Au cours de l’activité 1, vous êtes invité à lire le chapitre 11 du livre Fundamentals of ecological modelling de Jørgensen et Fath (2011), qui résume quelques approches de modélisation spatiale et présente quatre exemples de modèles spatiaux en écologie.
L’activité 2 consiste à lire le chapitre 8 du livre Modelling complex ecological dynamics, rédigé par Brecking, Pe’er et Matsinos (2011), qui se consacre exclusivement à la présentation du formalisme des automates cellulaires, une approche de modélisation spatiale simple en écologie.
L’activité 3 vous permet d’explorer la dynamique d’un automate cellulaire par la simulation du modèle NetLogo « Segregation », un modèle connu, conçu par l’économiste Schelling.
Finalement, au cours de l’activité 4, vous êtes convié à lire un article du biologiste Wooton qui décrit la méthodologie utilisée pour modéliser un système intertidal avec un automate cellulaire.
Introduction
La dimension spatiale est absente de la modélisation systémique à compartiments (module 2.2). En effet, cette approche de modélisation suppose que les compartiments, les processus au sein des compartiments et les flux entre les compartiments se déroulent au même endroit. Cette simplification facilite le développement de modèles conceptuels ainsi que la recherche de solutions mathématiques (Jørgensen et Fath, 2011). Bien qu’il soit raisonnable pour répondre à certaines questions de recherche de supposer cette absence de relations spatiales dans et entre les compartiments, les phénomènes écologiques et environnementaux sont intrinsèquement spatiaux. L’inclusion de la dimension spatiale est donc souvent requise.
Les modèles spatiaux permettent de représenter la distribution spatiale, le mouvement et la dispersion des constituants d’un système. Ils permettent aussi d’inclure la notion de proximité dans les interactions entre les constituants. Par exemple, un modèle de croissance des arbres pourrait inclure la distance entre les arbres puisqu’elle influence leur compétition pour la lumière. De plus, les modèles spatiaux permettent d’explorer l’effet de l’hétérogénéité spatiale sur l’évolution du système. Par exemple, la distribution des nutriments dans les sols est hétérogène et rend certaines zones plus productives que d’autres pour la croissance des plantes.
Depuis les années 70, différentes approches de représentation et de modélisation de l’espace ont été développées. L’avènement des systèmes d’information géographique et de la télédétection a contribué à offrir des outils plus sophistiqués de représentations de l’espace (Jørgensen et Fath, 2011). Les domaines scientifiques que sont l’écologie du paysage et de la modélisation spatiale ont pu bénéficier de ces avancées.
Passez maintenant à l’activité de lecture 1. Cette lecture est une introduction à la modélisation spatiale. Elle vous familiarise avec la terminologie et les concepts propres à la modélisation spatiale. Elle vous fait aussi connaître différentes approches et exemples de modèles qui intègrent la dimension spatiale.
Terminologie et concepts
L’espace peut être représenté sous forme de variable continue ou discrète. Lorsque l’espace est représenté de façon discrète, il est subdivisé en entités distinctes qui sont désignées par les coordonnées x, y et (ou) z. Ces coordonnées sont des nombres entiers positifs (c’est-à-dire 1, 2, 3, etc.). Lorsque l’espace est représenté de façon continue, il dépend des coordonnées, x, y, et (ou) z qui sont des nombres réels positifs. Ces coordonnées peuvent prendre un nombre infini de valeurs entre les entiers (c’est-à-dire 1,001, 1,0012, 1,00128, etc.). La figure 2.3.1 illustre des exemples de représentations continues (a, b, et c) et discrètes (d, e, f) de l’espace en une, deux et trois dimensions.

Figure 2.3.1Méthodes de représentation de l’espace de façon continue (a, b, et c) et discrète (d, e, et f). Figure adaptée de Thomas et Huggett (1980).
Deux concepts sont fondamentaux en modélisation spatiale : l’échelle et la résolution spatiale. L’échelle spatiale se définit comme l’étendue d’un processus écologique. La réponse des organismes vivants dans un écosystème, à différents processus écologiques, peut se manifester sur différentes échelles spatiales (figure 2.3.2). Plusieurs processus opèrent aussi à différentes échelles spatiales (Jørgensen et Fath, 2011). Le modélisateur doit ainsi faire un choix judicieux de l’échelle spatiale à utiliser qui est spécifique à sa question de recherche.
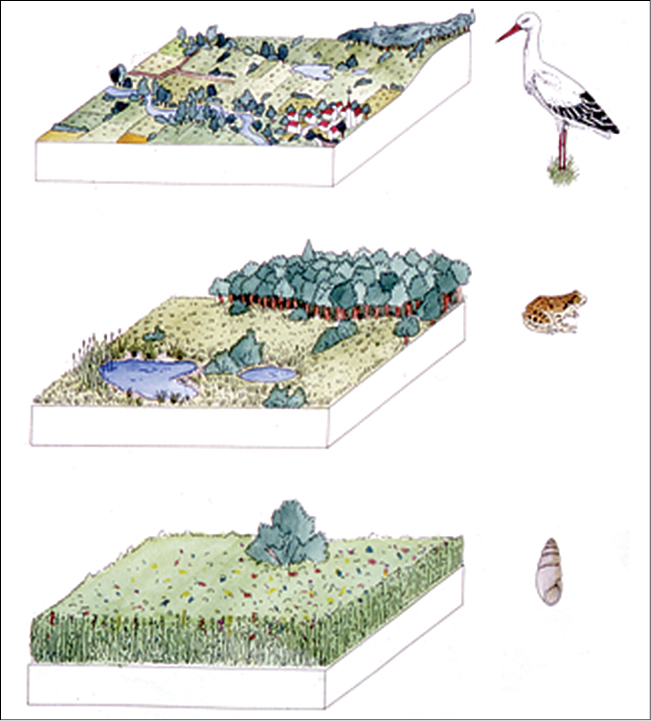
Figure 2.3.2 Différence d’échelles spatiales auxquelles les organismes opèrent. Figure parue dans Henle et al. (2014).
La résolution est, quant à elle, la plus petite unité spatiale à laquelle l’espace est représenté (figure 2.3.3). Son choix peut dépendre de la question de recherche, du système à modéliser ou des données disponibles. Par exemple, le modèle de l’exemple 1, présenté dans le chapitre 11 du livre Fundamentals of ecological modelling de Jørgensen et Fath, s’intéresse à la succession forestière de l’épinette de Norvège à la suite d’un chablis ayant affecté une superficie d’environ 128 ha. Dans cet exemple, la résolution spatiale choisie est petite (1 m2). Cependant, dans le modèle de l’exemple 2 qui étudie la succession à long terme de la savane sur une grande étendue (90,000 km2), la résolution est plus grande (3 km2).
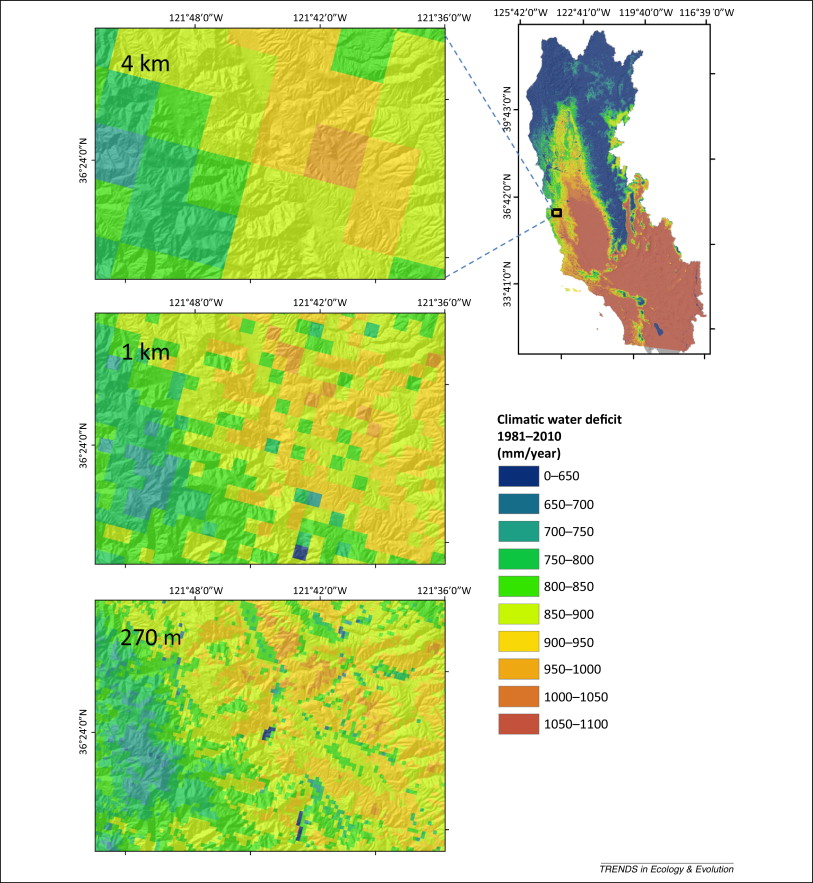
Figure 2.3.3 Trois résolutions spatiales différentes dans un modèle étudiant les changements de distributions spatiales des espèces causés par les changements climatiques. Figure parue dans Hannah et al. (2014).
Généralement, un modèle qui utilise une grande échelle spatiale et une fine résolution spatiale se traduit par des simulations exigeantes en temps de calcul. Parfois des compromis doivent être faits quant à la résolution et à l’échelle spatiale utilisées pour réduire le temps de calcul.
Approches de modélisation spatiale
Modèles de réaction-diffusion
Le chapitre 11 du livre de Jørgensen et Fath présente quelques-unes des premières approches de modélisation spatiale. Vous avez appris que certains modèles spatiaux sont basés sur la théorie de la réaction-diffusion. Ces modèles sont utilisés pour étudier les systèmes dans lesquels les composantes, par exemple des espèces animales ou végétales, interagissent et se dispersent localement sur le paysage. Ces modèles s’expriment par des équations spatiales différentielles composées de deux termes : un terme représentant la réaction et un terme représentant la diffusion des composantes.

L’équation (1) représente une équation de réaction-diffusion pour la variable N(x,y,t). Cette variable peut, par exemple, représenter la taille de la population d’une espèce donnée. Le premier terme, R(N,t), est la fonction de réaction et il modélise généralement la croissance ou la production de la variable N. Le second terme,  , modélise la diffusion de N dans l’espace. Le paramètre D est le taux de diffusion et l’opérateur
, modélise la diffusion de N dans l’espace. Le paramètre D est le taux de diffusion et l’opérateur  , appelé le laplacien, désigne l’opération
, appelé le laplacien, désigne l’opération  .
.
La figure 2.3.4 montre différents patrons spatiaux produits par un modèle de réaction-diffusion développé pour étudier la formation de patrons spatiaux de végétation dans un milieu où les précipitations sont limitées (von Hardenberg, Meron, Shachak et Zarmi, 2001). Les patrons formés par le modèle s’apparentent à ceux observés dans les régions désertiques.

Figure 2.3.4 Patrons spatiaux produits par un modèle de réaction-diffusion (a-c). Patrons formés par l’herbe Paspalum vaginatum, observés dans la région désertique du Negev (Israël) (d-f). La distance entre les patrons d’herbe est de l’ordre de 15 cm. Figure adaptée de von Hardenberg et al. (2001).
Le modèle s’exprime par des équations différentielles pour la densité de biomasse d’herbe h et la densité d’eau souterraine e :
(2)

Dans l’équation pour l’herbe, la fonction de réaction F est basée sur des processus de croissance et de mortalité de l’herbe qui peuvent dépendre de la densité d’eau. La dispersion des plantes est modélisée par le terme de diffusion  . Dans l’équation pour l’eau, la fonction de réaction G intègre le niveau de précipitation, l’évaporation et l’absorption par les plantes. Le deuxième terme modélise la diffusion hydraulique et le troisième terme le ruissellement de l’eau. Tous les deux sont dépendants de la hauteur de ruissellement z.
. Dans l’équation pour l’eau, la fonction de réaction G intègre le niveau de précipitation, l’évaporation et l’absorption par les plantes. Le deuxième terme modélise la diffusion hydraulique et le troisième terme le ruissellement de l’eau. Tous les deux sont dépendants de la hauteur de ruissellement z.
Dans les modèles de réaction-diffusion le temps et l’espace sont des variables continues. Lorsque l’espace et le temps sont des variables discrètes, les modèles basés sur la théorie de la réaction-diffusion sont appelés réseaux d’applications couplées (coupled-map lattice). L’espace est alors représenté par une grille bidimensionnelle formée de cellules carrées identiques dont les coordonnées sont données par les variables x et y, et le temps est composé d’itérations (t = 1, 2, 3, …). Nous reviendrons sur le concept de discrétisation de l’espace et du temps lorsque nous discuterons plus bas des modèles basés sur le formalisme des automates cellulaires.
Les réseaux d’applications couplées ont été abondamment utilisés en écologie pour étudier l’effet de l’espace sur la coexistence de deux espèces aux interactions antagonistes. Par exemple, des espèces entretenant des relations de compétition, de prédateur-proie ou d’hôte-parasitoïde. La figure 2.3.5 illustre des patrons spatiaux qui émergent de réseaux d’applications couplées représentant la dynamique de population de deux espèces.

Figure 2.3.5 Patrons spatiaux produits par des réseaux d’applications couplées. a) Modèle de compétition entre deux espèces (Solé, Bascompte et Valls, 1992). Les cellules sont noires si la densité d’une des deux espèces est supérieure à 0,5; elles sont blanches si la densité est inférieure au seuil 0,5. b) Modèle d’hôte-parasite (Hassell, Comins et May, 1991). Les cellules rouges indiquent une forte densité d’une des deux espèces et le bleu une faible densité.
Les prédictions des modèles spatiaux quant à la coexistence des espèces peuvent être plus réalistes que les modèles non spatiaux. En effet, les modèles spatiaux prédisent généralement la coexistence des deux espèces pour un plus large éventail de paramètres que les modèles n’intégrant pas la dimension spatiale et qui tendent à prédire l’extinction d’une des deux espèces.
Les modèles basés sur la théorie de la réaction-diffusion sont appropriés pour modéliser un système dans lequel le ou les composantes se dispersent de façon passive. Dans les systèmes pour lesquels les composantes sont des organismes vivants faisant des choix actifs de déplacement (pour chercher de la nourriture ou un abri ou pour fuir un prédateur), des mécanismes de dispersion plus complexes que la diffusion doivent être modélisés.
Intégration de l’espace dans la modélisation systémique à compartiments
Il est possible d’intégrer la dimension spatiale dans la modélisation systémique à compartiments. Cette approche nécessite de diviser l’étendue spatiale du système à modéliser en cellules. L’espace est alors traité comme une variable discrète. Chaque cellule devient un compartiment du modèle et est liée aux cellules adjacentes par des flux. Les flux peuvent symboliser des échanges d’eau, d’air, d’animaux, d’énergie et de biens et services économiques (Costanza et Voinov, 2004).
Les modèles spatiaux à compartiments comptent plusieurs échelles spatiales (Costanza et Voinov, 2004). L’échelle spatiale propre aux cellules dans lesquelles la dynamique interne se déroule et l’échelle du paysage où émerge la dynamique produite par les cellules connectées. Il est aussi possible de modéliser une échelle spatiale plus fine à l’intérieure de chaque cellule. Les dynamiques à différentes échelles spatiales opèrent aussi sur différentes échelles temporelles.
La figure 2.3.6 illustre conceptuellement la structure d’un modèle spatial à compartiments. Ce modèle d’occupation du territoire représente un paysage dans lequel l’état de chaque cellule évolue en fonction de sa dynamique interne et de ses échanges avec les cellules voisines.
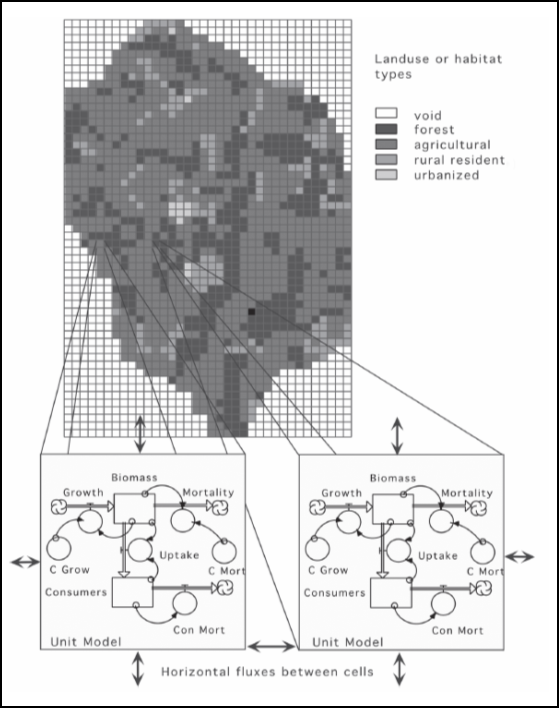
Figure 2.3.6 Modèle d’occupation du sol illustrant la structure générale des modèles spatiaux à compartiments. Figure reprise de Costanza et Voinov (2004).
L’approche de modélisation spatiale à compartiments est utile pour étudier des systèmes hydrodynamiques et atmosphériques. La résolution mathématique de ce type de modèle est complexe puisque des équations différentielles doivent être résolues pour chaque cellule représentée. La méthode des éléments finis est privilégiée pour résoudre numériquement ces équations. Dans ce cours, nous n’aborderons pas cette méthode d’analyse numérique qui requiert une formation avancée en mathématiques.
Le modèle CELSS de Sklar, Costanza et Day, présenté dans le chapitre 11 du livrede Jørgensen et Fath, utilise une approche de modélisation spatiale à compartiments pour étudier le changement d’habitats dans les terres humides côtières de la région marécageuse du delta d’Atchafalaya en Louisiane.
La zone marécageuse est divisée en plus d’un million de cellules d’une résolution de 50 m2. Chaque cellule constitue un modèle de simulation dynamique et non linéaire à deux compartiments, un compartiment pour les sédiments en suspension et l’autre pour les sédiments de fond. Chaque cellule est connectée aux cellules adjacentes et échange de l’eau, du sel et des sédiments avec ses voisines, rendant le modèle spatialement dynamique.
L’évolution du modèle permet d’explorer les changements d’habitats dans la zone marécageuse en déterminant l’état de chaque cellule (marais salé, marais d’eau douce, marais saumâtre, etc.) en fonction du niveau d’eau, de la salinité et de la profondeur des sédiments. La figure 2.3.7 illustre le territoire modélisé dans ses conditions initiales et plusieurs années plus tard suivant un scénario d’augmentation du niveau de la mer.
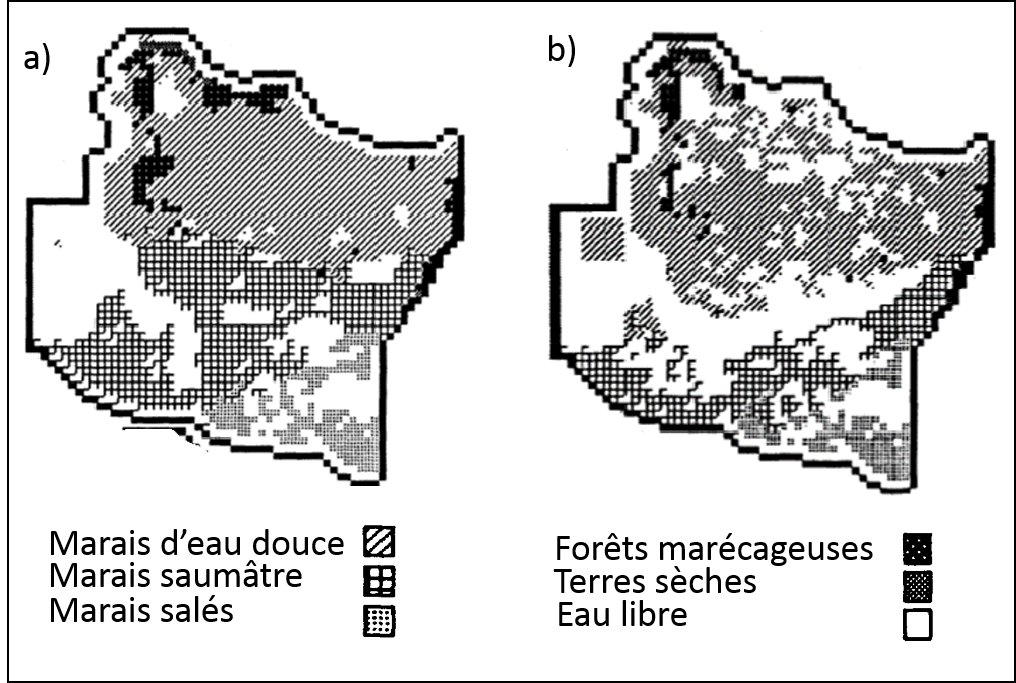
Figure 2.3.7 Prédictions du modèle spatial à compartiments CELSS. a) Le territoire aux conditions initiales (année 1978). b) Le territoire suivant la simulation d’un scénario d’augmentation du niveau de la mer (année 2033). Figure adaptée de Costanza, Sklar et White (1990) .
Automates cellulaires
Les automates cellulaires constituent une approche de modélisation spatiale basée sur la discrétisation de l’espace, du temps et des variables d’état. Les automates cellulaires forment un des plus simples formalismes de modélisation explicitement spatiale et leurs applications en écologie et en environnement sont nombreuses. C’est pour cette raison que, dans ce module, nous nous attardons plus longuement à cette approche de modélisation.
L’activité 2 consiste à lire un chapitre du livre rédigé par Breckling, Pe’er et Matsinos. Ce chapitre vous présente les principales caractéristiques des automates cellulaires et vous fournit des exemples d’application.
Propriétés principales
Dans un automate cellulaire, l’espace est discret. Il peut avoir une, deux ou trois dimensions. Généralement, en écologie et en environnement, les automates cellulaires ont deux dimensions. L’espace est divisé en cellules. Les cellules peuvent être de formes et de tailles identiques, généralement des carrés (figure 2.3.8 a). Dans ce cas, elles forment une grille et peuvent être désignées par leurs coordonnées cartésiennes. Les cellules peuvent aussi avoir des formes polygonales différentes propres aux structures du paysage qu’on cherche à modéliser, par exemple pour représenter des cadastres dans un territoire donné (figure 2.3.8 b).
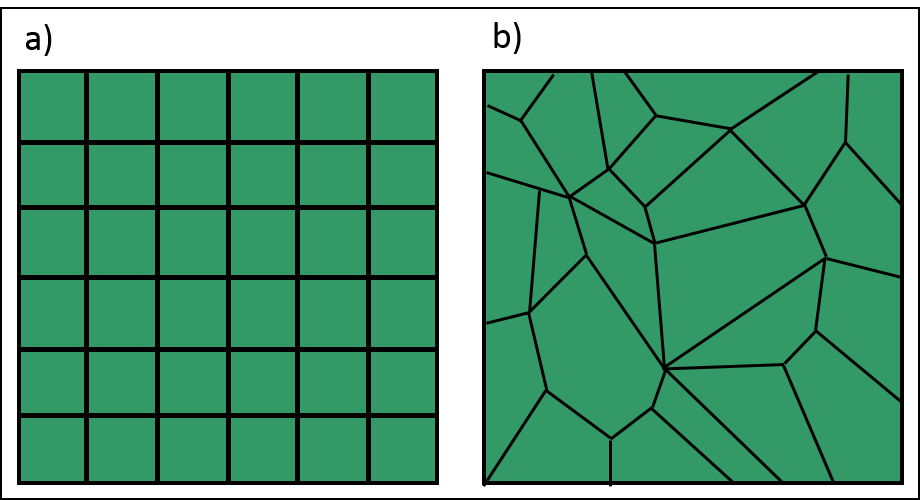
Figure 2.3.8 Exemples de discrétisation d’un espace bidimensionnel. a) Les cellules sont des carrés identiques. b) Les cellules sont des polygones de formes et de tailles variables.
Le temps est aussi une variable discrète dans les automates cellulaires. Ainsi, la simulation d’un automate cellulaire procède par itération de la variable t qui prend des valeurs entières (t = 1, 2, 3, …).
Finalement, les cellules d’un automate cellulaire sont décrites par une ou plusieurs variables d’état qui sont aussi discrètes. Une variable d’état est discrète lorsqu’elle peut prendre une valeur parmi un ensemble de valeurs distinctes. Ceci est différent d’une variable continue qui peut prendre un nombre infini de valeurs réelles. Par exemple, dans un automate cellulaire modélisant la dynamique de population d’une espèce donnée, une cellule peut être dans l’état 1 si elle est occupée par l’espèce modélisée, et dans l’état 0 si elle est vide. L’état d’une cellule est donc une variable discrète prenant les valeurs 0 ou 1.
Cette discrétisation des états différencie les automates cellulaires des réseaux d’applications couplées. Dans un réseau d’applications couplées modélisant la dynamique d’une population, l’état de chaque cellule serait plutôt modélisé par la densité de population, prenant ainsi des valeurs réelles positives.
Considérons maintenant un automate cellulaire représentant un territoire forestier. La figure 2.3.9 présente ce territoire par une grille composée d’un ensemble de cellules. Les cellules peuvent être dans quatre états différents. Elles peuvent être occupées par un plan d’eau, composées de jeune forêt, composées de forêt mature, ou en proie à un feu de forêt.
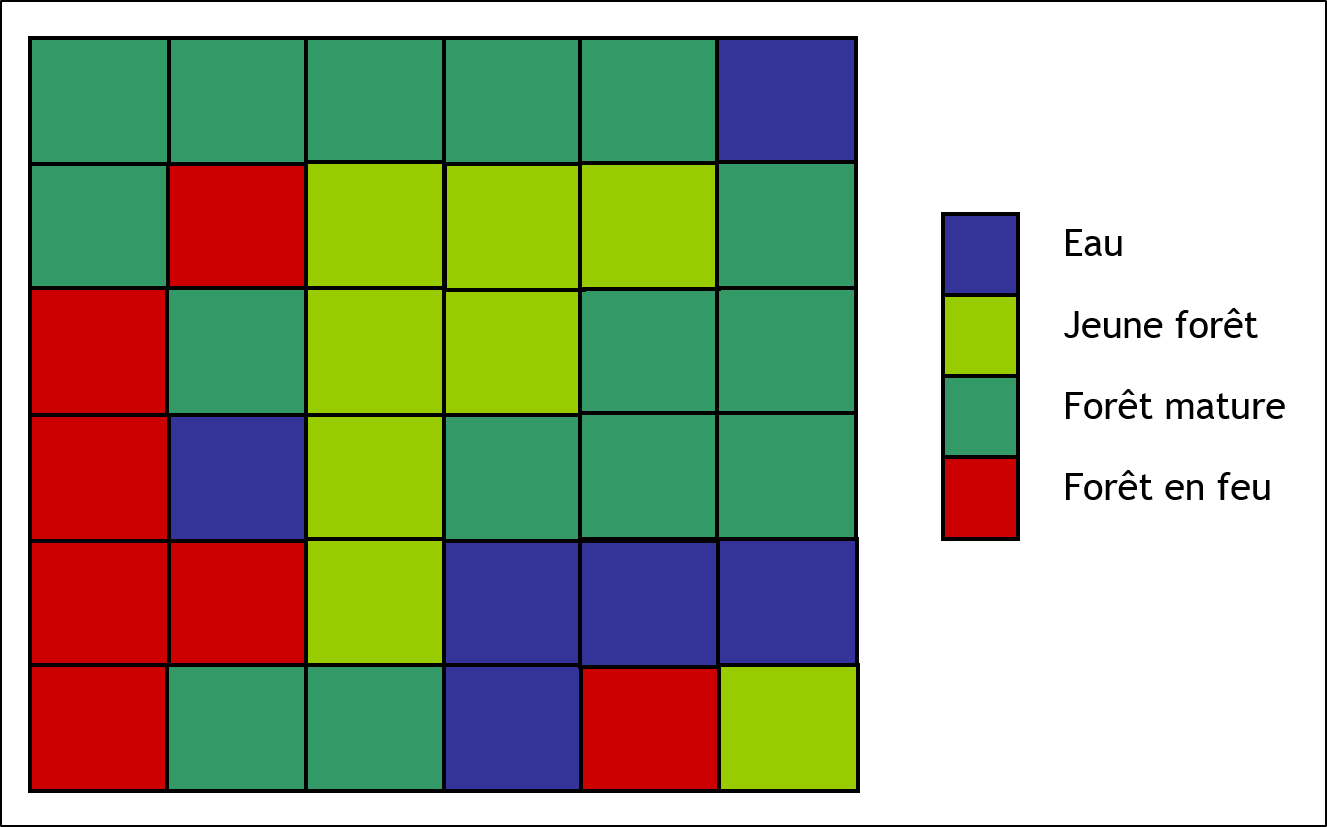
Figure 2.3.9 Automate cellulaire représentant un territoire forestier.
Les cellules peuvent être dans quatre états : eau (bleue), jeune forêt (vert pâle), forêt mature (vert foncé) et forêt en feu (rouge).
L’évolution d’un automate cellulaire est menée par des règles de transition qui contrôlent le changement d’état des cellules à chaque itération. Les changements d’état des cellules sont généralement effectués de façon synchrone : toutes les cellules subissent une transition en même temps. Dans le cas du territoire forestier, ces règles de transition pourraient être les suivantes :
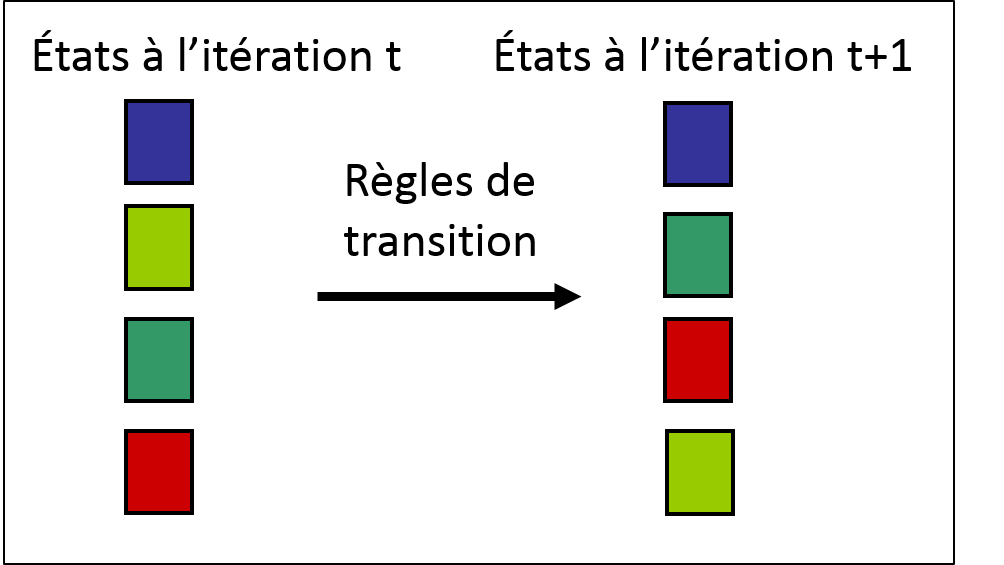
Figure 2.3.10 Règles fictives pour un automate cellulaire représentant un territoire forestier.
Les états possibles sont les suivants : eau (bleue), jeune forêt (vert pâle), forêt mature (vert foncé) et forêt en feu (rouge).
Cependant, les automates cellulaires sont des modèles explicitement spatiaux qui cherchent à modéliser les relations spatiales entre les cellules. Ainsi, les règles de transition dans un automate cellulaire tiennent compte non seulement de l’état d’une cellule, mais aussi de l’état des cellules qui lui sont adjacentes. Un concept important des automates cellules est donc la notion de proximité spatiale ou de voisinage.
Il existe plusieurs définitions de voisinage d’une cellule. Le type de voisinage choisi dépend du processus spatial qu’on cherche à modéliser. Deux définitions classiques sont le voisinage de von Neumann (contenant la cellule ciblée par la transition et les cellules au nord, au sud, à l’est et à l’ouest) et le voisinage de Moore (qui inclut le voisinage de von Neumann et les cellules au nord-est, au nord-ouest, au sud-est et au sud-ouest). La figure 8.2 du chapitre 8 du livre Modelling complex ecological dynamics de Breckling, Pe’er et Matsinos illustre ces deux types de voisinage.
Les règles de transition pour le territoire forestier données à la figure 2.3.10 peuvent ainsi être corrigées en tenant compte du voisinage de von Neumann. La figure 2.3.11 procure un exemple de règles de transition.
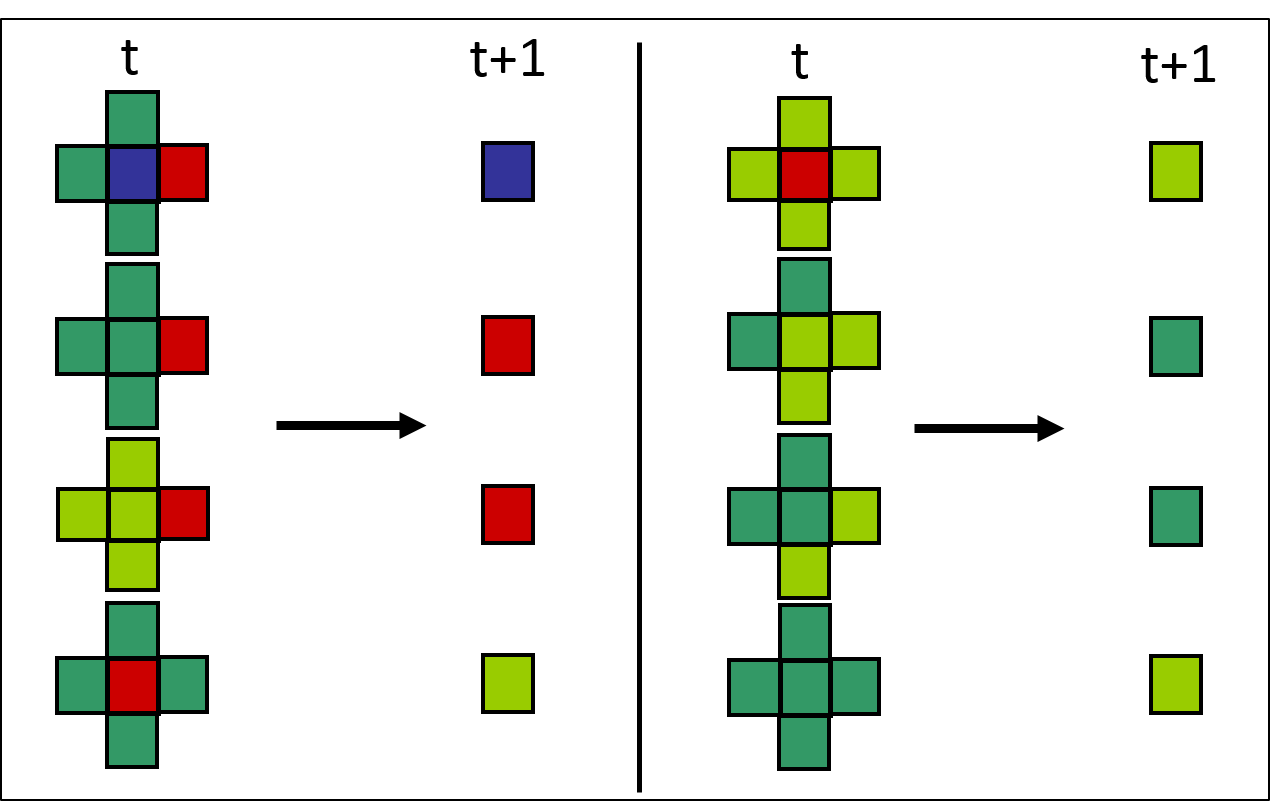
Figure 2.3.11 Règles de transition en fonction du voisinage de von Neumann pour l’automate cellulaire représentant un territoire forestier.
Les états possibles sont les suivants : eau (bleue), jeune forêt (vert pâle), forêt mature (vert foncé) et forêt en feu (rouge).
Les règles de transition peuvent être déterministes ou encore probabilistes (figure 2.3.12). Lorsque les règles sont probabilistes, le changement d’état d’une cellule est fonction d’une probabilité définie préalablement.
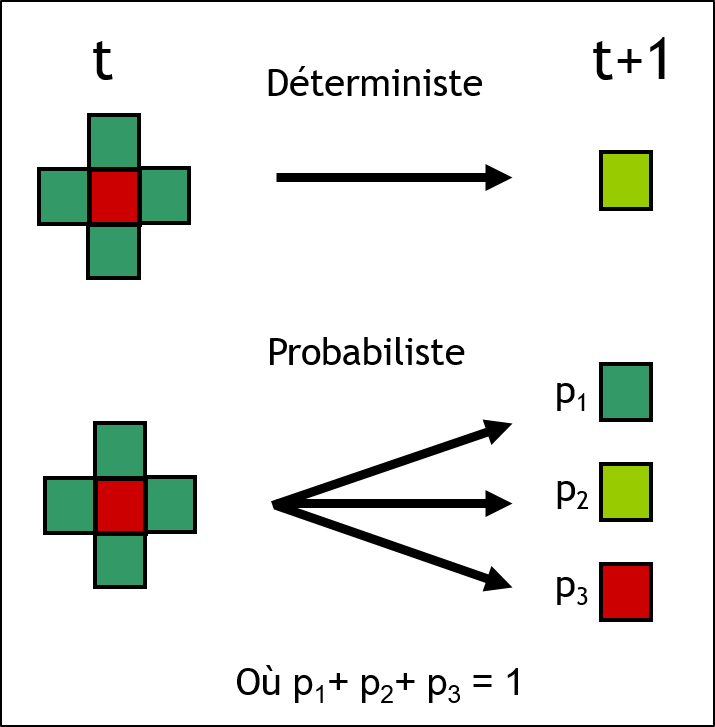
Figure 2.3.12 Règles de transitions déterministes ou probabilistes d’un automate cellulaire.
Les états possibles sont les suivants : eau (bleue), jeune forêt (vert pâle), forêt mature (vert foncé) et forêt en feu (rouge).
Lors de la conception d’un automate cellulaire, il importe de définir les conditions frontières de la grille. En effet, les règles de transition s’appliquent à toutes les cellules de la grille, y compris celles situées en bordure de celle-ci dont le voisinage est incomplet (figure 2.3.13 a). La notion de voisinage est étendue aux cellules frontières en définissant une zone tampon formée de cellules entourant la grille (figure 2.3.13 b). L’analyse de la dynamique d’un automate cellulaire exclut la zone tampon et considère seulement les cellules de la grille.
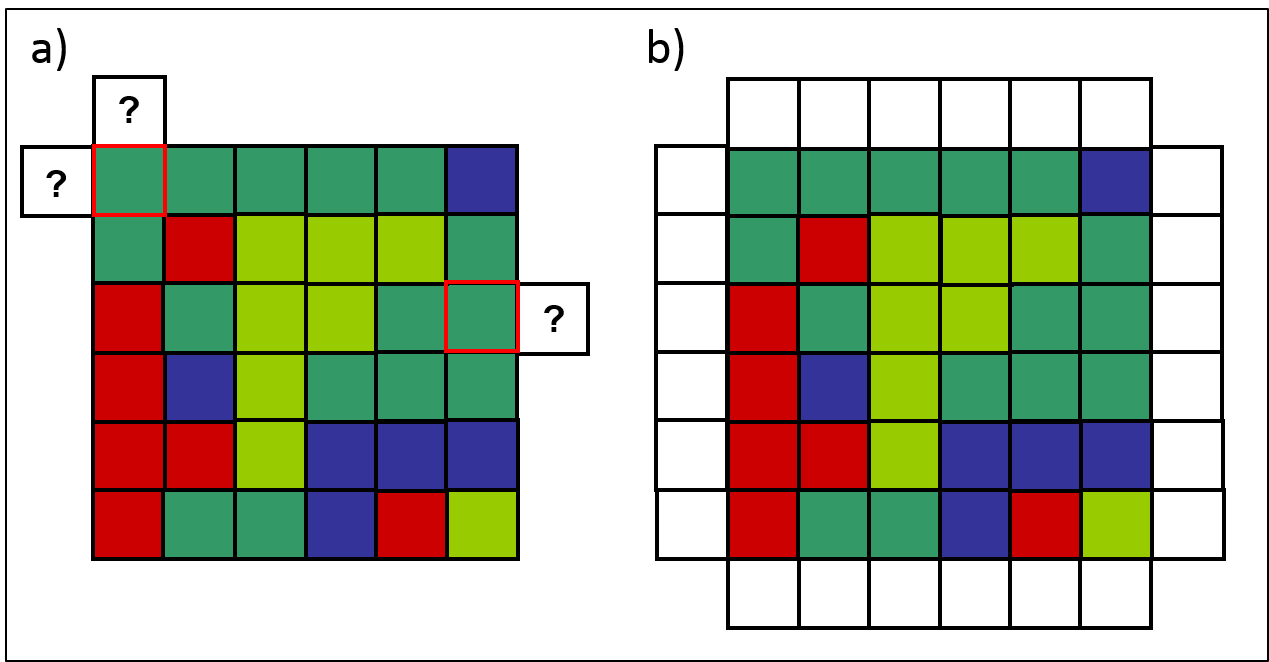
Figure 2.3.13 Les conditions frontières dans l’automate cellulaire représentant un territoire forestier. a) Les cellules en bordure de la grille ont un voisinage incomplet (ici de von Neumann). b) Les conditions frontières sont définies en créant une zone tampon qui entoure la grille de l’automate cellulaire.
Les cellules de cette zone tampon peuvent prendre différentes valeurs. On parle de conditions frontières à valeur fixe (ou absorbante) lorsque les cellules de la zone tampon prennent une valeur identique et fixe qui n’évoluera pas au cours de la simulation (figure 2.3.14 a). Les conditions frontières sont dites réfléchissantes lorsque chaque cellule de la zone tampon prend la même valeur que la cellule qui lui est directement adjacente (figure 2.3.14 b). Finalement, les conditions frontières sont dites périodiques lorsque chaque cellule de la zone tampon prend la même valeur que la cellule qui lui est située à l’opposé sur la même ligne ou colonne (figure 2.3.14 c). Les conditions frontières périodiques peuvent être illustrées par un tore comme le présente la figure 8.3 du chapitre 8 du livre de Breckling, Pe’er et Matsinos. Dans les deux derniers cas, la valeur des cellules dans la zone tampon est dynamique et évolue au cours de la simulation.
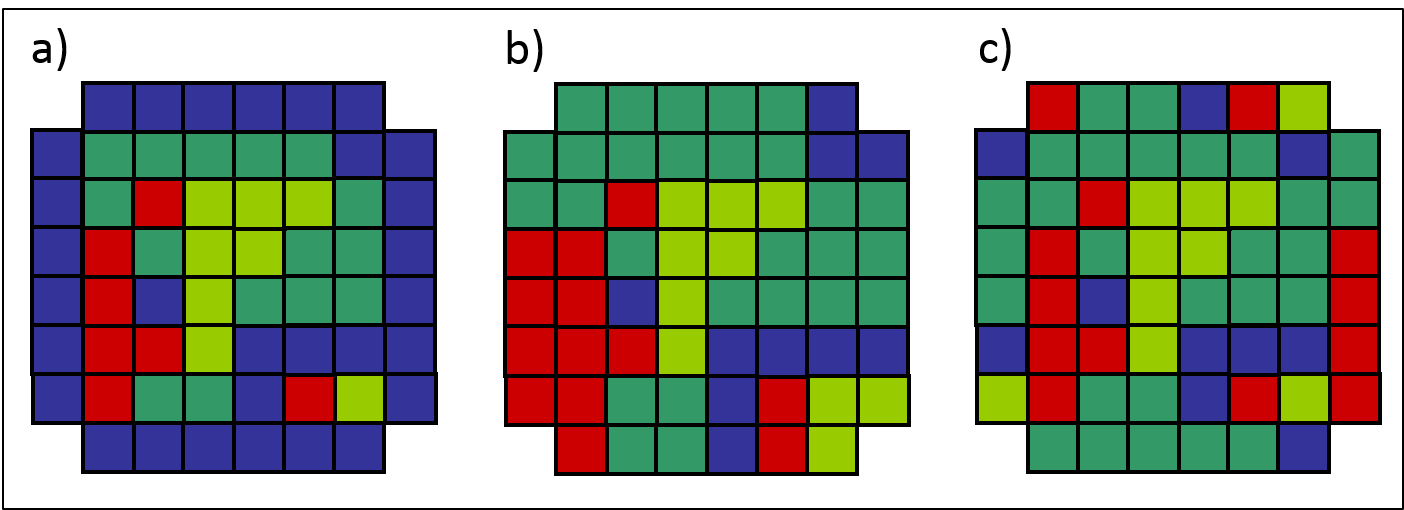
Figure 2.3.14 Les trois types de conditions frontières dans l’automate cellulaire du territoire forestier. a) Conditions frontières à valeur fixe. b) Conditions frontières réfléchissantes. c) Conditions frontières périodiques.
Origines
Le chapitre 8 du livre de Breckling, Pe’er et Matsinos vous renseigne sur l’origine des automates cellulaires. Celle-ci remonte aux années 50. Ce sont les mathématiciens John von Neumann et Stanislaw Ulam, intéressés à simuler des patrons spatiaux capables de s’autoreproduire, qui ont développé ce formalisme. Par autoreproduction, on entend la faculté d’un patron spatial de produire, par voie de simulation informatique, un autre patron spatial sans que soit nécessaire l’intervention du modélisateur.
Les automates cellulaires ont été popularisés, dans les années 60, par le mathématicien John Conway qui créa le jeu informatique appelé le jeu de la vie (the game of life). Dans ce jeu, les cellules placées sur une grille bidimensionnelle peuvent prendre deux états : « mort » ou « vivant ». Les règles simples de transition de l’état des cellules de mort à vivant, et l’inverse, donne lieu à des patrons hautement complexes. Des dynamiques stationnaires, oscillatoires et même d’autoreproduction peuvent émerger.
La section 8.3 du chapitre 8 du livre de Breckling, Pe’er et Matsinos vous explique comment fonctionne le jeu de la vie. Les curieux peuvent aussi explorer le modèle du jeu de la vie dans le répertoire de modèles disponibles dans NetLogo (voir le menu File > Models Library > Sample Models > Computer Science > Cellular Automata > Life).
Le physicien Stephen Wolfram a contribué de façon importante à la compréhension de la dynamique produite par les automates cellulaires en étudiant de façon systématique les automates cellulaires à une dimension avec deux variables d’état. La figure 2.3.15 présente trois exemples d’automates cellulaires étudiés par Wolfram.
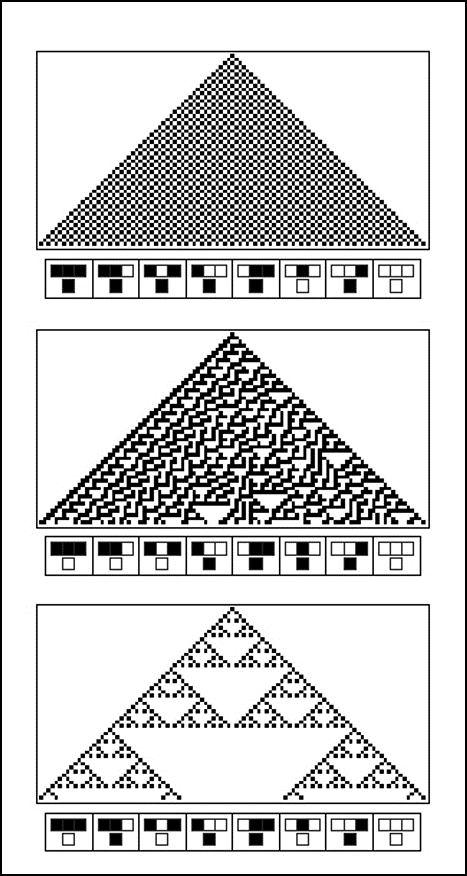
Figure 2.3.15 Exemples d’automates cellulaires étudiés par Wolfram. Figure adaptée de Wolfram (2002).
Notez que ces automates cellulaires ont une seule dimension. Ils forment une ligne horizontale composée de cellules. Les cellules peuvent avoir deux états : blanc ou noir. Initialement, l’automate cellulaire est dans l’état présenté par la ligne supérieure : seule la cellule au centre de la ligne est noire, toutes les autres sont blanches. Les règles de transition sont données au bas de chaque image. Ces règles sont fonction de l’état de la cellule ciblée et l’état des cellules à droite et à gauche de cette dernière. À chaque itération, l’état de chaque cellule est mise à jour et une nouvelle ligne horizontale est ajoutée sous la ligne précédente.
En étudiant les patrons formés par ces automates cellulaires, Wolfram a déterminé que quatre classes de dynamique étaient possibles :
a) la formation de structures homogènes;
b) la formation de structures stables ou périodiques (qui se répètent à intervalle régulier);
c) la formation de structures chaotiques ou apériodiques (qui ne semblent pas se répéter);
d) la formation de structures localisées complexes, qui semblent être à mi-chemin entre des structures périodiques et apériodiques, et qui peuvent se propager.
Des exemples de ces classes de dynamique sont illustrés à la figure 2.3.16 avec la simulation d’un automate cellulaire à une dimension et deux états (blanc ou noir). La simulation a montre que les cellules évoluent vers l’état blanc et forment ainsi une structure homogène. La simulation b illustre la formation d’une structure stable. Il est alors possible de prédire l’état futur de l’automate cellulaire. La simulation c illustre la formation d’une structure apériodique. Dans ce cas, il est impossible de prédire l’évolution de l’automate cellulaire. Finalement, la simulation d illustre la formation d’une structure complexe localisée. Cette structure évolue pendant plusieurs itérations et chevauche des structures stables et périodiques.

Figure 2.3.16 Exemples des quatre classes de dynamiques selon Wolfram. a) Dynamique homogène. b) Dynamique stable. c) Dynamique apériodique ou chaotique. d) Dynamique complexe. Figure adaptée de Wolfram (1984).
Les automates cellulaires sont des modèles fascinants puisque, malgré les règles de transition fort simple qui s’appliquent localement aux cellules, ils peuvent produire une gamme diversifiée de dynamiques qui émergent à l’échelle de la grille.
Les automates cellulaires se sont révélés des outils fort appropriés pour modéliser des processus spatiaux en écologie et en environnement. Par exemple, des automates cellulaires ont servi à modéliser la dynamique de plantes (Silvertown, Holtier, Johnson et Dale, 1992), les feux de forêt (Malamud, Morein et Turcotte, 1998), la propagation d’épidémies (Sirakoulis, Karafyllidis et Thanailakis, 2000), ou encore le changement d’utilisation du sol (White et Engelen, 1993).
Les sections qui suivent vous présentent deux exemples de modélisation par automate cellulaire.
Exemple 1 : le modèle de ségrégation de Schilling
Thomas Schelling est un économiste reconnu pour ses contributions au domaine de la théorie des jeux. La théorie des jeux est l’étude des stratégies de prise de décision à l’aide de modèles mathématiques. Les travaux de recherche de Schelling ont permis une meilleure compréhension des stratégies menant à des conflits et à la coopération. Il s’est vu octroyer le prix Nobel d’économie en 2005 pour ces travaux sur ce thème.
Le modèle de ségrégation de Schilling a été développé dans les années 70 pour étudier le phénomène de ségrégation ethnique, observé dans les grandes villes américaines (Schelling, 1971). Examinez par exemple la figure 2.3.17 qui illustre ce phénomène dans la ville de New York en 2000. Schilling s’est intéressé à comprendre les facteurs qui pouvaient mener à ce phénomène. Outre les facteurs socio-économiques et politiques, il s’est demandé si le comportement des individus, en l’occurrence leur décision de résider dans un quartier plutôt qu’un autre, pouvait contribuer à générer ces patrons spatiaux de ségrégation.
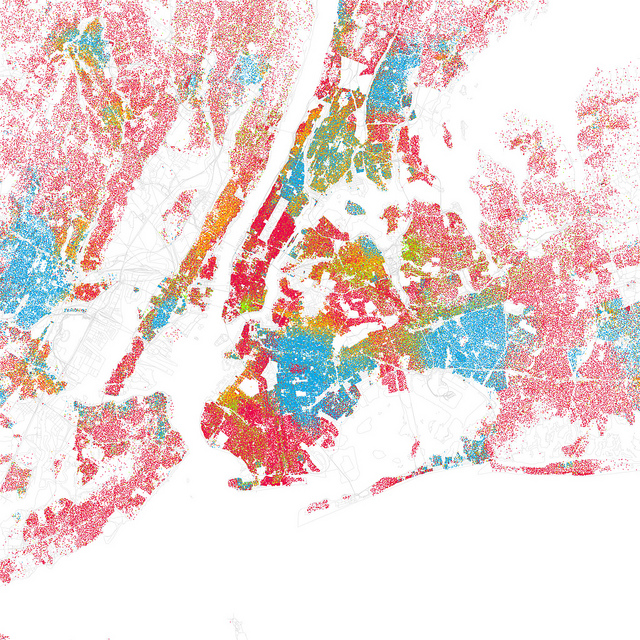
Figure 2.3.17 Le phénomène de ségrégation ethnique dans la ville de New York. © Eric Fischer.
Cette carte a été créée à partir des données d’un recensement fait en 2000. Les points rouges sont des adresses occupées par des Caucasiens, les bleus par des Afro-Américains, les jaunes par des Latino-Américains et les verts par des Asiatiques.
L’activité 3 vous invite à explorer le modèle de ségrégation de Schilling. Ce modèle est un automate cellulaire. Il vous permet d’observer que des décisions prises individuellement par des résidents peuvent générer une ségrégation. Ainsi, un patron observé à une grande échelle spatiale, comme celle d’une ville, peut être le résultat de comportements se déroulant à une petite échelle spatiale, comme celle d’un voisinage.
Exemple 2 : modélisation par automate cellulaire d’un système intertidal
Le modèle de Schilling constitue un exemple théorique d’un automate cellulaire. Il permet certes d’améliorer notre compréhension des processus qui peuvent mener à la ségrégation, mais les règles de transition ne sont pas conçues à partir de données empiriques, ce qui restreint le réalisme du modèle. L’activité suivante vous convie à la lecture d’un article sur le développement d’un automate cellulaire conçu à partir de données empiriques pour étudier la dynamique d’une communauté d’espèces de la zone intertidale. Un milieu intertidal est l’espace du littoral sujet à la marée.
Dans les travaux de recherche ayant mené à la publication de cet article, Wootton s’est questionné sur l’effet des perturbations par les vagues et des interactions locales entre les individus de diverses espèces de la zone intertidale sur la dynamique de la communauté.
Dans une communauté d’espèces de la zone intertidale, les moules tendent à envahir la roche nue et les individus adjacents appartenant à d’autres espèces. Elles se fixent au couvert et s’attachent entre elles grâce à leurs filaments byssaux. Les vagues ont pour effet d’extraire les moules du couvert. Ces perturbations se propagent aux moules voisines puisque celles-ci sont connectées entre elles. Ce mécanisme génère des ouvertures de tailles variables sur la roche. Ces patrons spatiaux se caractérisent par une distribution de fréquence de la taille des ouvertures qui suit une loi de puissance.
Les espèces intertidales sont peu mobiles et de dimensions faciles à manipuler. De plus, la dynamique de la communauté se déroule sur une échelle de temps relativement rapide. Pour ces raisons, l’écosystème intertidal se prête bien à la modélisation par automate cellulaire.
Wootton a étudié 14 sites dans la zone intertidale de l’île Tatoosh dans l’État de Washington pendant six ans. La figure 2.3.18 illustre le design expérimental utilisé par Wootton. Chaque site est constitué d’une grille de 60 x 60 cm divisée en cellules de 6 x 6 cm. L’état de chaque cellule est recensé au printemps et à l’automne pendant la durée de l’étude. Il y a 15 états possibles : roche nue, trois tailles de moules et 11 autres espèces intertidales dont différentes espèces d’algues et de bernacles. La présence de perturbations est aussi recensée. Les changements d’état des cellules entre deux inventaires permettent de concevoir des règles de transition probabilistes pour l’automate cellulaire.

Figure 2.3.18 Design expérimental utilisé par Wootton pour le développement d’un automate cellulaire d’un écosystème intertidal. © Wootton.
L’automate cellulaire développé par Wootton est constitué d’une grille de 100 x 300 cellules aux conditions frontières absorbantes. Il utilise un voisinage de von Neumann. Chaque cellule peut être dans un des 15 états répertoriés. Le modèle est simulé pendant 500 itérations au cours desquelles des perturbations sont initiées et l’état de chaque cellule est actualisé selon les règles de transitions. Ces règles dépendent de l’état de la cellule, de la présence d’une moule dans une cellule voisine ainsi que de sa taille et si la cellule a subit une perturbation à l’itération précédente.
Wootton a simulé plusieurs variantes de ce modèle (voir Box 1 dans l’article) :
- l’automate cellulaire complet tel que décrit plus haut, un modèle n’incluant pas les perturbations (local interactions model);
- un modèle n’incluant pas les perturbations et ne considérant pas les cellules voisines dans les règles de transition (Markov chain model);
- un modèle ignorant les espèces autres que les moules (detailed mussel/no mussel model);
- un modèle basé seulement sur la présence ou l’absence de moules sans détail quant à leur taille (basic mussel/no mussel model).
Il a comparé la composition taxonomique et la distribution de fréquence de la taille des ouvertures produites par ces modèles à celles observées empiriquement.
Wootton démontre que tous les modèles qui incluent les multiples espèces prédisent correctement la composition taxonomique d’une communauté intertidale (voir la figure 4 de l’article). Ce résultat implique qu’une étude cherchant à prédire la composition taxonomique ne requière pas le développement d’un modèle détaillé qui simule les interactions locales et les perturbations.
Wootton démontre cependant que les distributions de la taille des ouvertures varient grandement entre les modèles (voir la figure 2 de l’article). Ce résultat signifie que la modélisation des interactions locales entre les cellules à l’aide des règles de transition dépendantes du voisinage et des perturbations, est importante pour reproduire les patrons spatiaux observés dans les communautés intertidales.
Le concept d’auto-organisation est souvent mentionné dans l’article de Wootton. L’auto-organisation est le processus spontané par lequel un patron ordonné ou une structure cohérente émerge dans un système sans nécessiter la contribution d’un agent ou d’une force externe. L’auto-organisation est un thème récurrent dans les modèles d’automates cellulaires puisque les règles de transitions fondées sur le voisinage peuvent produire des patrons spatiaux ordonnés. Ce thème est aussi fréquent dans les modèles orientés-individus et multi-agents qui sont le sujet du prochain module (module 2.4).
Références
Breckling, B., Pe’er, G. et Matsinos, Y. G. (2011). Cellular automata in ecological modelling. Dans F. Jopp, H. Reuter et B. Breckling (dir.), Modelling complex ecological dynamics (p. 105-117). Berlin : Springer.
Costanza, R., Sklar, F. H. et White, M. L. (1990). Modeling coastal landscape dynamics. Bioscience, 40(2), 91-107.
Costanza, R. et Voinov, A. (2004). Introduction: Spatially explicit landscape simulation models. Dans R. Costanza et A. Voinov (dir.), Landscape Simulation Modeling (p. 3-20). New York : Springer
Fischer, E. [Figure]. Repéré à www.flickr.com/photos/walkingsf/4981444199/sizes/z/in/set-72157624812674967/. Licence : https://creativecommons.org/licenses/by-sa/4.0/
Hannah, L., Flint, L., Syphard, A. D., Moritz, M. A., Buckley, L. B. et McCullough, I. M. (2014). Fine-grain modeling of species’ response to climate change: Holdouts, stepping-stones, and microrefugia. Trends in Ecology & Evolution, 29(7), 390-397. doi : https://dx.doi.org/10.1016/j.tree.2014.04.006
Hassell, M. P., Comins, H. N. et May, R. M. (1991). Spatial structure and chaos in insect population dynamics. Nature, 353(6341), 255-258.
Henle, K., Potts, S., Kunin, W., Matsinos, Y. G., Simila, J., Pantis, J., . . . Settele, J. (dir.). (2014). Scaling in ecology and biodiversity conservation : Advanced Books.
Jørgensen, S. E. et Fath, B. D. (2011). Spatial modelling. Dans Fundamentals of ecological modelling (p. 347-368). Oxford : Elsevier.
Malamud, B. D., Morein, G. et Turcotte, D. L. (1998). Forest fires: An example of self-organized critical behavior. Science, 281(5384), 1840-1842.
Schelling, T. C. (1971). Dynamic models of segregation. Journal of Mathematical Sociology, 1(2), 143-186.
Silvertown, J., Holtier, S., Johnson, J. et Dale, P. (1992). Cellular automaton models of interspecific competition for space – the effect of pattern on process. Journal of Ecology, 80(3), 527-534.
Sirakoulis, G. C., Karafyllidis, I. et Thanailakis, A. (2000). A cellular automaton model for the effects of population movement and vaccination on epidemic propagation. Ecological Modelling, 133(3), 209-223. doi : 10.1016/s0304-3800(00)00294-5
Solé, R. V., Bascompte, J. et Valls, J. (1992). Nonequilibrium dynamics in lattice ecosystems: Chaotic stability and dissipative structures. Chaos: An Interdisciplinary Journal of Nonlinear Science, 2(3), 387-395. doi : https://dx.doi.org/10.1063/1.165881
Thomas, R. W. et Huggett, R. J. (1980). Modelling in geography: A mathematical approach. London : Harper & Row.
von Hardenberg, J., Meron, E., Shachak, M. et Zarmi, Y. (2001). Diversity of vegetation patterns and desertification. Physical Review Letters, 87(19). doi : 19810110.1103/PhysRevLett.87.198101
White, R. et Engelen, G. (1993). Cellular-automata and fractal urban form : A cellular modeling approach to the evolution of urban land-use patterns. Environment and Planning A, 25(8), 1175-1199. doi : 10.1068/a251175
Wolfram, S. (1984). Universality and complexity in cellular automata. Physica D: Nonlinear Phenomena, 10(1–2), 1-35. doi : https://dx.doi.org/10.1016/0167-2789(84)90245-8.
Wolfram, S. (2002). A new kind of science. Champaign, IL. : Wolfram Media, Inc.
Wootton, J. T. [Photographie]. Repéré à https://woottonlab.uchicago.edu/?page_id=133. Utilisé avec la permission de l’auteur.
Wootton, J. T. (2001). Local interactions predict large-scale pattern in empirically derived cellular automata. Nature, 413(6858), 841-844.