Module 2.1 Évolution conceptuelle et historique de la modélisation écologique
Résumé du module
Dans ce module, nous étudions le contexte scientifique dans lequel la modélisation écologique s’est établie comme domaine de recherche. Quelles sont les avancées marquantes ayant permis de transformer l’écologie, une science autrefois purement qualitative, en science quantitative où la modélisation est considérée comme pouvant fournir des contributions importantes à la compréhension du fonctionnement des écosystèmes?
Au cours de ce module, vous ferez la lecture du chapitre 3 du livre Modelling complex ecological dynamics de Brecking, Jopp et Reuter. Ce chapitre résume les étapes marquantes de l’évolution conceptuelle et historique de la modélisation écologique ainsi que son importance pour l’écologie moderne.
Introduction
La modélisation écologique a vu le jour dans les années 1920 et elle constitue maintenant un domaine de recherche bien établi. La modélisation écologique repose sur une quantification de l’écosystème étudié – de ses composantes et des processus qui les relient. Comme tout domaine de recherche, celui de la modélisation écologique est en évolution constante. Les paradigmes changent, les modèles se perfectionnent, les applications se multiplient.
L’émergence et l’évolution de la modélisation écologique sont étroitement liées aux avancées produites dans d’autres domaines de recherche, que ce soit en mathématiques, en physique, en informatique ou autre. Par exemple, l’accès à des ordinateurs de plus en plus puissants permet de modéliser des processus beaucoup plus complexes par voie de simulation.
Pour saisir l’état des lieux en modélisation écologique, nous commençons cette partie du cours par la lecture du chapitre 3 du livre de Brecking, Jopp et Reuter, qui résume les étapes marquantes de l’évolution de ce domaine. Ce chapitre vous apprend qui sont les précurseurs de la modélisation écologique, les pionniers et les acteurs qui ont contribué à son essor en diversifiant les approches et les applications.
Cette synthèse ne fournit pas un portrait complet de tous les acteurs ayant contribué au développement de la modélisation écologique, mais permet tout de même de mieux comprendre les origines des différentes approches de modélisation qui seront étudiées dans les prochains modules.
Les précurseurs de la modélisation écologique
L’écologie est une discipline qui découle de l’histoire naturelle, une science largement descriptive et qualitative. Ainsi, l’initiative de quantifier les relations entre l’homme et son environnement émerge davantage de disciplines quantitatives comme la physique, les mathématiques, l’économie et la démographie.
Malthus
Robert Malthus (1766 – 1834) est un penseur et un académicien ayant fortement marqué les domaines de l’économie politique et de la démographie. En 1798, il publie un essai intitulé An essay on the principle of population,qui traite des causes et des conséquences de la croissance de la population humaine. Malthus avait observé que, dans la nature, seule survit une faible proportion des descendants produits par la reproduction des plantes et des animaux. L’homme, selon lui, était aussi sujet à la surreproduction en l’absence de processus de contrôle de sa population. Si la fertilité de chaque individu est identique dans le temps et dans l’espace, le taux de croissance de la population est constant. Il quantifie ainsi la croissance de la population humaine par un processus exponentiel.
La croissance exponentielle d’une population est représentée à la figure 2.1.1. L’image de gauche illustre la croissance d’une population où chaque individu donne naissance à deux individus au cours de sa vie. L’imagine centrale démontre comment une population de taille initiale n0 augmente d’un facteur de deux à chaque génération. Finalement, la figure de droite illustre l’évolution temporelle de la population n suivant une fonction exponentielle de taux constant égal à deux.
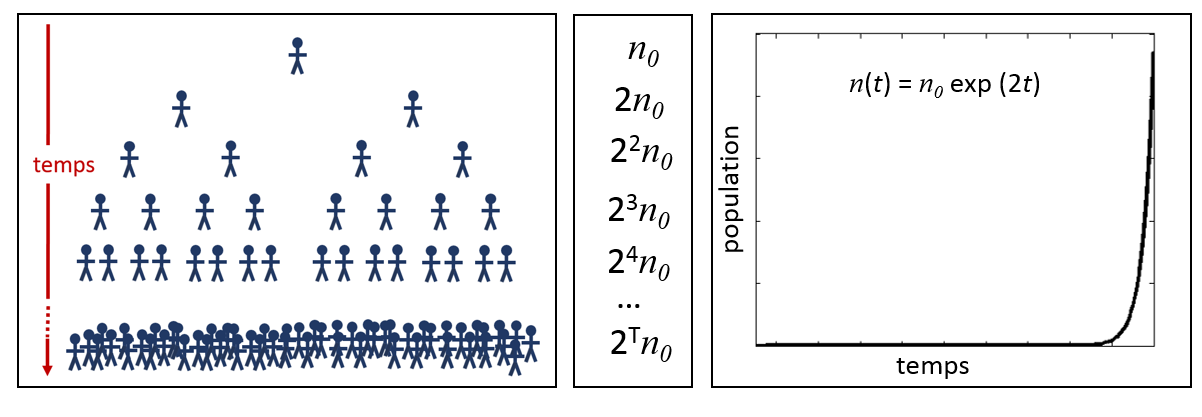
Figure 2.1.1 Croissance exponentielle de la population.
Selon Malthus, les ressources naturelles nécessaires à la survie de la population n’augmentent pas suffisamment rapidement pour satisfaire cette croissance exponentielle de la population, ce qui mènera éventuellement à la famine, à la propagation de maladies, et à la misère. Ce phénomène s’appelle la catastrophe malthusienne. Cette vision diverge de celle qui suggère que le progrès lié à l’augmentation de la productivité et à l’établissement de structures sociales permet d’éviter une telle catastrophe. La contribution de Malthus est toujours au cœur des débats portant sur l’utilisation accrue par l’homme des ressources environnementales limitées.
Verlhulst
Le mathématicien Pierre François Verlhulst (1804 – 1849), en réponse au modèle de Malthus, a proposé un modèle selon lequel un accroissement illimité de la population est impossible. Ce modèle porte le nom de modèle logistique et est toujours couramment utilisé en écologie des populations. Dans ce modèle, le taux de croissance n’est pas une constante, comme cela est le cas dans le modèle de Malthus. Le taux de croissance est une fonction qui dépend de la taille de la population et de la capacité limite du milieu. La capacité limite, aussi appelée la capacité de soutien ou la capacité de charge, est la taille maximale de la population qu’un milieu donné peut supporter.
La figure 2.1.2 illustre la croissance logistique d’une population. La taille de la population est exprimée par la variable n. La croissance de la population est donnée par la fonction r qui dépend de n et de la capacité limite exprimée par le paramètre K. Lorsque la taille de la population est inférieure à la capacité de soutien (représentée par la ligne rouge), la population croît (courbe pointillée). La population cesse de croître lorsque sa taille atteint la capacité de soutien. Cependant, lorsque la taille de la population est supérieure à la capacité de soutien, celle-ci décroît jusqu’à atteindre la capacité de soutien (courbe tiretée).
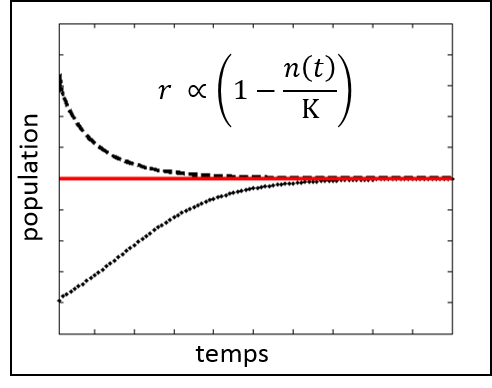
Figure 2.1.2 Croissance logistique de la population.
Les modèles de Malthus et Verlhulst correspondent aux premières initiatives de quantification de processus observés en écologie, notamment en dynamique des populations, par des équations mathématiques. Cet exercice de quantification qui a pour objectif de vérifier des hypothèses et de faire des prédictions correspond aux fondements mêmes de la modélisation.
Les pionniers de la modélisation écologique
La science au début du 20e siècle est marquée par les avancées en physique, particulièrement la théorie de la relativité d’Einstein et la découverte de la mécanique quantique. L’importance d’expliquer des phénomènes observés par une théorisation et une formulation mathématique se manifeste aussi en écologie.
Initialement, les écologistes tentent de décrire comment certaines variables évoluent dans le temps en utilisant des équations différentielles simples. Une équation différentielle est une équation mathématique qui relie une fonction d’une ou de plusieurs variables à ses dérivées. Par exemple, considérons une variable donnée x. La dérivée de x, dont l’expression mathématique est dx/dt, correspond à la variation de x sur un intervalle de temps infinitésimalement petit. Supposons que cette variation soit connue et s’exprime par la fonction F(x,t). L’équation différentielle pour x prend la forme dx/dt = F(x,t). Le calcul différentiel et intégral permet alors de résoudre cette équation et de déterminer x(t). Ainsi, il devient possible de savoir comment la variable x évolue dans le temps.
Il n’est pas nécessaire de bien connaître le calcul différentiel et intégral pour suivre ce cours. Ne vous laissez pas impressionner par les équations mathématiques! Retenez seulement que c’est un outil mathématique utile pour résoudre des problèmes qui cherchent à déterminer comment une variable évolue dans le temps ou dans l’espace.
Les équations (1) présentent, respectivement, les équations différentielles des modèles de Malthus et de Verlhulst. L’évolution temporelle de la population n est déterminée par des équations liant son accroissement au taux de croissance r, à la capacité limite de l’environnement K et à sa taille initiale n0. Les règles du calcul différentiel et intégral permettent de trouver la solution d’une équation différentielle si elle est suffisamment simple.
(1)

Lotka et Volterra
Alfred Lotka (1880 – 1949) et Vito Volterra (1860 – 1940) ont développé indépendamment l’un de l’autre, mais à la même époque, des équations décrivant une dynamique simple entre un prédateur et sa proie. Alors que les modèles de Malthus et Verlhulst représentent la dynamique d’une seule population, le modèle de Lotka-Volterra décrit pour la première fois la dynamique qui résulte de l’interaction entre deux populations.
Le modèle de Lotka-Volterra est fondé sur l’hypothèse selon laquelle deux organismes interagissent de façon isolée dans un environnement homogène et constant. Ce sont des conditions théoriques difficilement observables puisque les écosystèmes, bien entendu, sont constitués d’une multitude d’organismes en interaction sujets à des variations environnementales. Cependant, ce modèle constitue une assise fondamentale sur laquelle de nombreuses variantes améliorant son réalisme et son applicabilité ont été développées et le sont toujours. Aujourd’hui, l’étude du modèle de Lotka-Volterra est un élément incontournable des cours d’écologie des populations (Roughgarden, 1979). Nous verrons ce modèle plus en détail dans le module 2.2.
La simplicité du modèle de Lotka-Volterra permet de résoudre ses équations explicitement à l’aide du calcul différentiel et intégral. Cependant, les variantes plus complexes du modèle, notamment celles décrivant les interactions entre plusieurs populations, doivent être résolues de façon approximative en utilisant des méthodes numériques. Ces méthodes nécessitent des connaissances avancées en mathématiques, ce qui a retardé l’étude de modèles plus complexes en écologie. Aujourd’hui, l’utilisation d’ordinateurs permet aux modélisateurs de résoudre ces modèles complexes.
Bertalanffy
Ludwig von Bertalanffy (1901 – 1972) est un biologiste et philosophe reconnu pour ses contributions à la théorie des systèmes, popularisée notamment, en 1968, par la publication de son livre General system theory. La théorie des systèmes, développée dans les années 1950, unit des principes et des concepts interdisciplinaires dans le but d’étudier le fonctionnement général des systèmes. Plusieurs chercheurs provenant de disciplines aussi variées que la biologie, la linguistique, et la sociologie ont contribué au développement de cette théorie qui a permis d’établir des ponts entre des sphères de connaissances qui évoluaient jusqu’alors de manière isolée.
La théorie des systèmes propose une vision holistique des systèmes. Elle va à l’encontre d’une approche réductionniste et soutient qu’on ne peut pas comprendre le fonctionnement d’un système en simplifiant son étude à quelques composantes ou sur des échelles de temps et d’espace limitées.
La théorie des systèmes met l’accent sur les interactions entre les composantes d’un système. Un système peut alors être décrit par un réseau complexe de composantes interconnectées. La recherche fondée sur la théorie des systèmes s’intéresse à comprendre la relation entre la connectivité des composantes d’un système et la dynamique globale qui en émerge. Les concepts de boucles de rétroaction, d’auto-organisation et d’émergence sont propres à la théorie des systèmes et seront discutés à plusieurs moments dans cette partie du cours.
Bertalanffy suggère aussi l’importance de considérer les systèmes biologiques comme des systèmes ouverts sujets à des flux changeants d’énergie et de matière. Les modèles conventionnels qui découlent de la physique classique sont appropriés pour décrire des systèmes fermés étudiés à l’état d’équilibre dans les conditions contrôlées d’un laboratoire, mais sont inadéquats pour expliquer la majorité des systèmes vivants.
Lindeman, et les frères Odum
Le chapitre 3 du livre de Brecking, Jopp et Reuter, que vous avez lu plus tôt, présente quelques chercheurs ayant contribué à mettre en vigueur une perspective quantitative de l’écologie.
Parmi ces chercheurs figure Raymond Lindeman (1915 – 1942) qui a initié le concept de dynamique trophique en écologie. Depuis, il est maintenant commun de représenter un écosystème par son réseau trophique, c’est-à-dire un modèle décrivant les relations trophiques (qui-mangent-qui) entre les espèces ou les groupes fonctionnels. Un réseau trophique est un ensemble de chaînes alimentaires. Plusieurs niveaux trophiques y sont représentés : les producteurs, les consommateurs et les décomposeurs. La dynamique trophique repose sur le principe qu’il existe un transfert d’énergie et de biomasse entre les niveaux trophiques.
La figure 2.1.3 illustre le réseau trophique d’un écosystème aquatique et terrestre. La figure 2.1.4 illustre, quant à elle, le réseau trophique associé au sapin baumier (balsam fir, en anglais, sur la figure). La complexité de ce réseau est beaucoup plus grande. La tordeuse de bourgeons de l’épinette (Choristoneura fumiferana) ainsi que cinq autres herbivores se nourrissent du sapin et sont eux-mêmes les hôtes de 66 différents parasitoïdes. Si vous désirez en savoir plus sur cet insecte défoliateur, vous pouvez consulter le site web du ministère des Forêts, de la Faune et des Parcs du Québec.
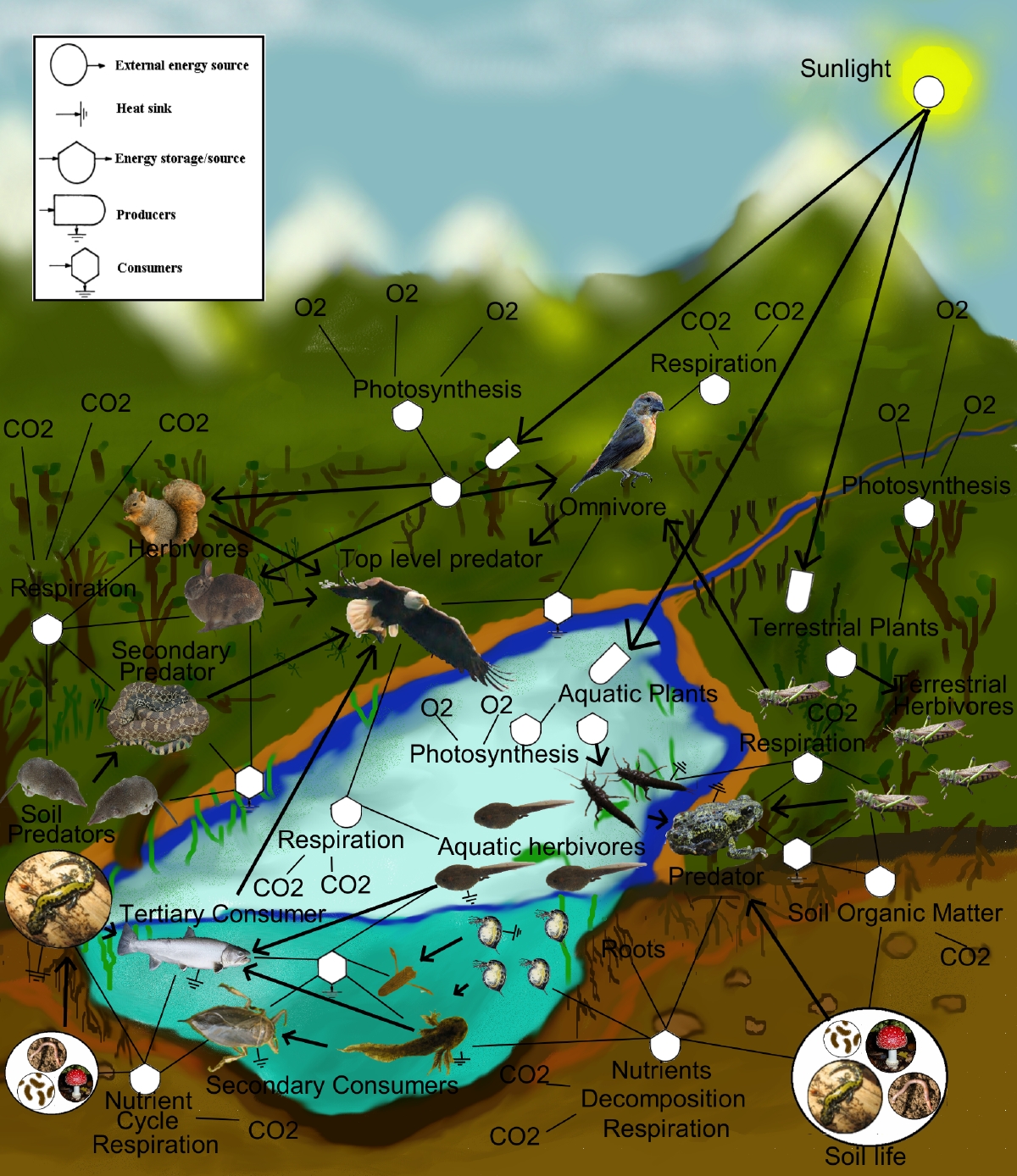
Figure 2.1.3 Réseau trophique d’un écosystème terrestre et aquatique (Thompsma, 2010).
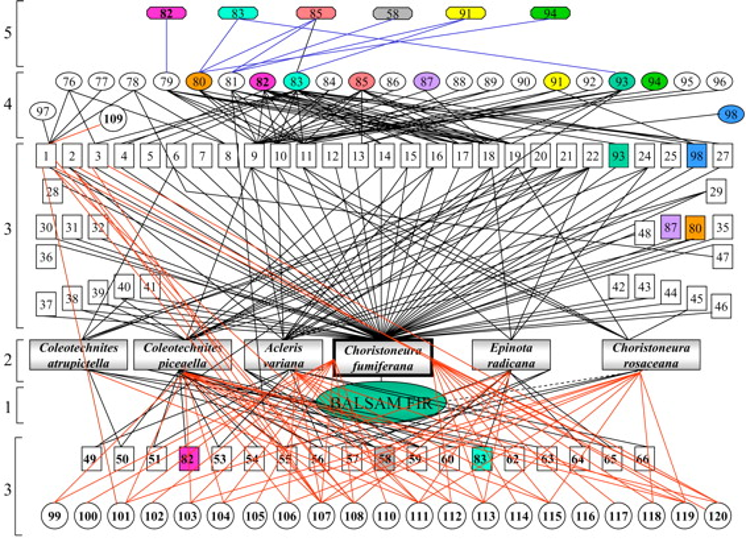
Figure 2.1.4 Le réseau trophique du sapin baumier. Figure parue dans Eveleigh et al. 2007.
Six espèces d’herbivores (rectangles) se nourrissent du sapin baumier (ellipse verte). Ces herbivores sont les hôtes de 66 parasitoïdes primaires (carrés), 21 pathogènes primaires (cercles), 23 parasitoïdes secondaires (ovales), un pathogène secondaire et six parasitoïdes tertiaires (octogones).
Comprendre le fonctionnement des réseaux trophiques est un sujet de recherche substantiel en écologie. Les modèles de réseaux trophiques permettent, entre autres, d’étudier comment les forces ascendantes (provenant des niveaux inférieurs du réseau) et descendantes (provenant des niveaux supérieurs) régulent la dynamique trophique et influencent la structure de communautés multitrophiques.
Les frères Odum ont joué un rôle fondamental dans l’application de la théorie des systèmes en écologie. Eugene Odum (1913 – 2002) est l’auteur du célèbre manuel Fundamentals of ecology (Odum, 1953). Ce livre procure une vue d’ensemble complète des principes et des concepts de l’écologie et de l’environnement selon une approche systémique mettant l’accent sur les relations quantitatives au sein de ces systèmes. E. Odum a influencé des générations d’écologistes en popularisant le concept d’écosystème et en contribuant à établir l’écologie comme une discipline à part entière.
Howard Odum (1924 – 2002) a émis l’idée de représenter les flux énergétiques au sein des écosystèmes en utilisant l’analogie des circuits électriques. Avec ses collègues, H. Odum a développé une forme de langage, appelé Energy systems language, permettant de décrire ces circuits. La figure 2.1.5 illustre les symboles propres à ce langage.
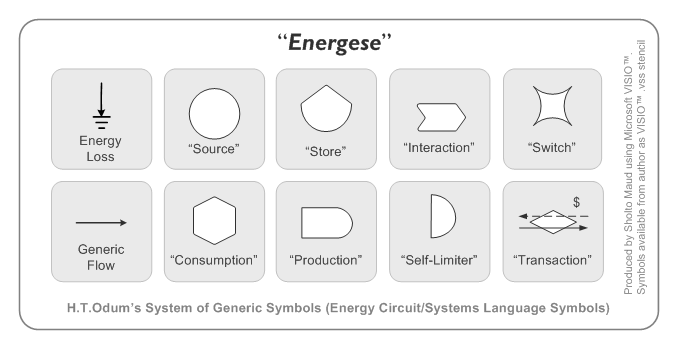
Figure 2.1.5 Symboles développés par H. Odum et ses collègues pour représenter les flux énergétiques dans les écosystèmes (Sholto Maud, 2007).
Cette approche permet une représentation conceptuelle, sous forme de diagramme, des échanges d’énergie entre les composantes d’un écosystème. La figure 2.1.3 utilise ce formalisme pour illustrer un réseau trophique. Nous avons appris au module 1.2 que l’élaboration d’un modèle conceptuel est une étape importante du processus de modélisation. De plus, le formalisme développé par H. Odum facilite la traduction des échanges d’énergie représentés sous forme d’équations mathématiques.
Forrester, Meadows, Patten, Jørgensen
Durant la deuxième partie du 20e siècle, des chercheurs de disciplines variées appliquent l’approche systémique pour développer des modèles propres à leur système d’étude. Cette approche se nomme modélisation systémique et sera présentée au module 2.2.
Jay Forrester (1918 –), un ingénieur informatique, est une figure marquante en dynamique des systèmes. Il utilise une approche systémique pour modéliser les cycles économiques dans le milieu industriel (1961) ainsi que la dynamique urbaine (1969). Son approche de modélisation est éventuellement généralisée dans une perspective de développement durable. En effet, le Club de Rome, un groupe de réflexion (think tank, en anglais), fondé en 1968, formé d’experts intéressés aux questions de politique internationale, demande à Forrester de concevoir un modèle permettant d’évaluer la durabilité des ressources naturelles à l’échelle mondiale.
Sous la direction de Forrester, un groupe de scientifiques, dont Donnella Meadows, Dennis Meadows et Jørgen Randers, développe le modèle World2 qui devient éventuellement le modèle World3. La figure 2.1.6 illustre le modèle conceptuel de World2. Cette figure est tirée du livre World dynamics (1971) de Forrester et donne un aperçu de la complexité du modèle.
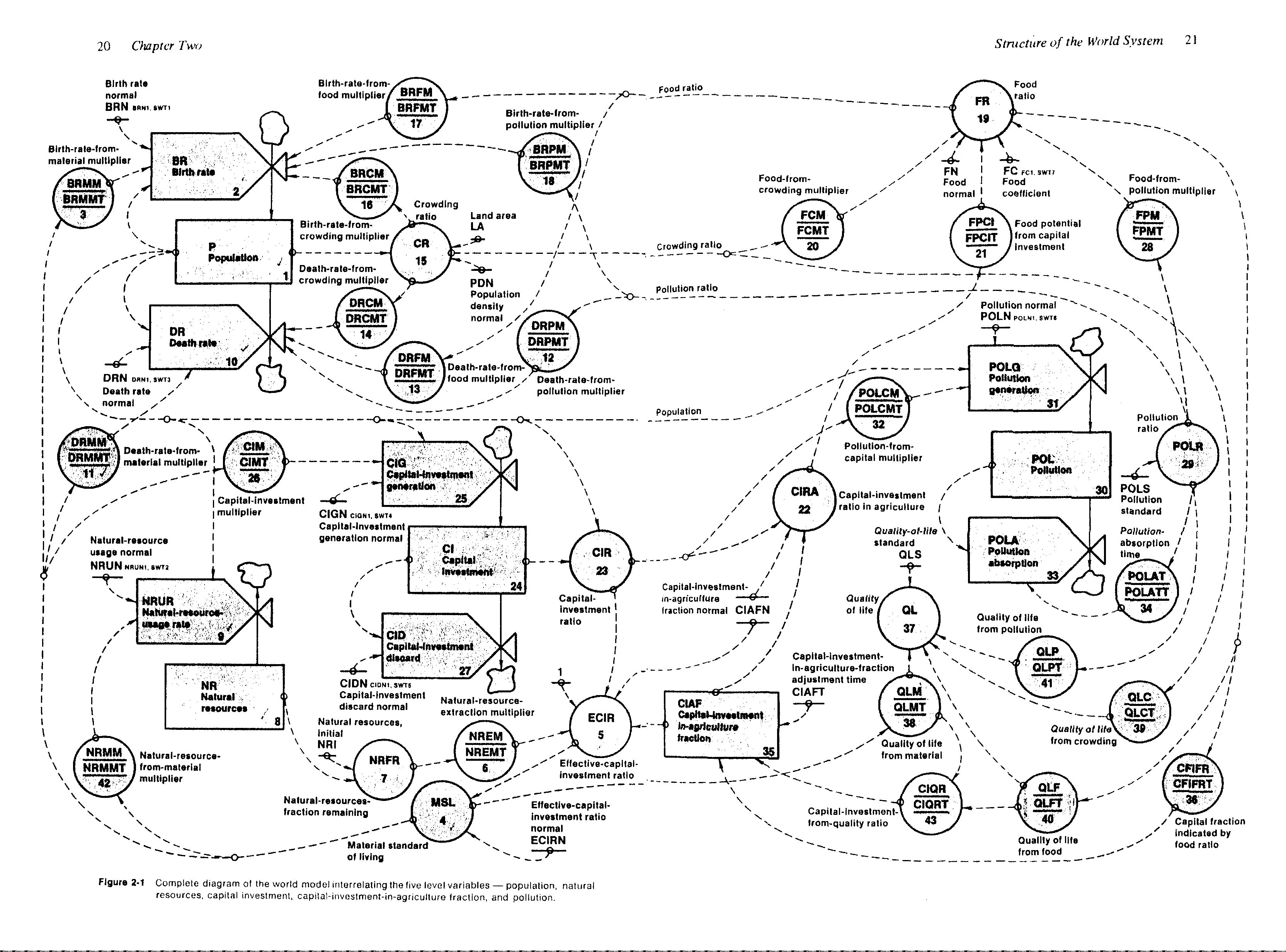
Figure 2.1.6 Modèle conceptuel de World2. Figure parue dans World dynamics (1971) de Forrester.
Les résultats pessimistes prédits par ce modèle ont reçu considérablement d’attention de la part du public lors de la publication du rapport Limits to growth (Meadows, Meadow, Randers et Behrens III, 1972). Nous étudierons plus en détail ce modèle dans le module 3.1.
L’écologiste Bernard Patten, quant à lui, démontre l’applicabilité de l’approche systémique en écologie en publiant Systems analysis and simulation in ecology (1971) . Ce livre présente de nombreux exemples de modèles d’écosystèmes terrestres et constitue un premier ouvrage sur la modélisation écologique.
L’écologiste Sven Erik Jørgensen (1934 –) a grandement contribué à la reconnaissance de la modélisation écologique comme une discipline à part entière. Par exemple, en 1975, il fonde la revue scientifique Ecological Modelling, offrant ainsi un forum spécialisé aux écologistes pratiquant la modélisation. En 1986, il rédige le manuel Fundamentals of ecological modelling, qui compte maintenant plusieurs rééditions. Cet ouvrage est un incontournable pour l’apprentissage de la modélisation en écologie. Plusieurs activités de lecture proposées dans ce cours sont tirées de ce manuel. De plus, Jorgensen fonde la société internationale pour la modélisation écologie (International Society for Ecological Modelling) qui promeut les applications d’une approche systémique et la simulation en écologie et en gestion des ressources naturelles. Cette société finance, entre autres, des congrès, des symposiums et des formations en modélisation.
Les nouveaux paradigmes au cœur de l’essor de la modélisation écologique
Au cours des années 80, les approches de modélisation ont commencé à se diversifier. Il devenait impératif d’adopter des modèles plus adéquats pour représenter la dynamique de variables hétérogènes. En effet, les relations écologiques sont complexes et varient dans l’espace et dans le temps. Or, les modèles fondés sur des équations différentielles peuvent difficilement reproduire la complexité et l’hétérogénéité inhérentes aux systèmes écologiques et environnementaux.
Le paradigme orienté-objet
Le paradigme orienté-objet constitue un changement conceptuel en programmation informatique. Il a été élaboré dans les années 60 par les informaticiens Norvégiens Ole Johann Dahl (1931 – 2002) et Kristen Nygaard (1926 – 2002). La programmation informatique est la rédaction d’un code dans un langage informatique donné qui, une fois exécuté, permet de résoudre un problème. Les programmes informatiques traditionnels sont fondés sur le concept de la décomposition fonctionnelle, c’est-à-dire que le problème à résoudre est décomposé en plusieurs petits sous-problèmes pouvant être facilement résolus. Dans ce paradigme traditionnel, il existe un programme principal qui dirige la succession d’étapes nécessaires à la résolution des sous-problèmes. Ceci se nomme la programmation procédurale. Cependant, cette façon de faire crée des programmes qui sont très rigides. En particulier, un changement mineur au problème à résoudre peut entraîner des modifications majeures au programme principal.
Le paradigme orienté-objet permet de concevoir des programmes plus modulables. Ce paradigme est fondé sur le concept d’objet. Un objet est un type de données abstraites qui peut représenter une idée ou une entité réelle. Un objet est décrit par son état et son comportement. En l’occurrence, les objets peuvent interagir entre eux. La programmation orientée-objet utilise le comportement des objets et leurs interactions entre eux pour résoudre les problèmes. Ce paradigme repose, entre autres, sur la notion d’encapsulation, c’est-à-dire que les données propres à un objet peuvent être cachées du reste du programme.
L’élaboration de la programmation orientée-objet a ouvert la porte à de nouvelles options en modélisation écologique. En effet, ce paradigme s’applique bien à la résolution de problèmes en écologie et en environnement. D’abord, il y a une correspondance commode qui peut s’établir entre les objets du programme et les entités ou organismes du monde réel. De plus, ce formalisme se prête bien à la description de relations écologiques entre les objets (interactions interspécifiques, proximité spatiale, relations génétiques, etc.). Ainsi, la programmation orientée-objet permet plus aisément de résoudre des problèmes portant sur l’influence de la structure et de l’organisation au sein d’un écosystème sur sa dynamique.
La modélisation orientée-individu
La modélisation orientée-individu est un changement de paradigme par rapport à la modélisation systémique. La modélisation systémique sera présentée en détail dans le module suivant (module 2.2). Elle consiste à représenter un écosystème par un ensemble de compartiments liés entre eux par des flux de matière ou d’énergie. Par exemple, la figure 2.1.3 illustre différents compartiments (les herbivores, les omnivores, les grands prédateurs, etc.) et les échanges de biomasse entre ces derniers.
La modélisation orientée-individu consiste à représenter un écosystème par les entités mêmes qui composent le système. Nous verrons cette approche dans le module 2.4. Dans ce cas, une espèce donnée n’est pas représentée par un compartiment, mais par l’ensemble des individus qui forment sa population. Ce paradigme permet d’intégrer une plus grande variabilité au sein de l’écosystème modélisé puisque chaque individu peut être représenté par des caractéristiques distinctes.
Le paradigme orienté-objet est particulièrement approprié pour programmer des modèles orientés-individu. Les objets du programme peuvent facilement correspondre aux individus du modèle. Les premiers à avoir adopté ce paradigme de programmation en modélisation écologique sont les biologistes Heinrich Kaiser et Paulien Hogeweg (1943 –). Notez qu’il est aussi possible de programmer des modèles simples orientés-individu par une programmation procédurale.
L’auto-organisation
L’auto-organisation est le processus spontané par lequel une organisation ou une structure ordonnée émerge dans un système sans l’aide d’un agent ou d’une force extérieure. L’auto-organisation est causée par les interactions entre les composantes d’un système qui produisent des boucles de rétroaction (communément appelées feedback). Une boucle de rétroaction se produit lorsqu’une composante subit le contrecoup d’une interaction qu’elle a portée à une autre composante.
Le concept d’auto-organisation a initialement été révélé en physique et en chimie, notamment par les travaux d’Ilya Prigogine (1917 – 2003) qui lui ont valu le prix Nobel de chimie en 1977. En modélisation écologique, les modèles orientés-individus permettent de générer des processus auto-organisés. Nous reviendrons sur ce concept lorsque nous discuterons des automates cellulaires au module 2.3 et des modèles orientés-individus au module 2.4.
Références
Breckling, B., Jopp, F. et Reuter, H. (2011). Historical background of ecological modelling and its importance for modern ecology. Dans F. Jopp, H. Reuter et B. Breckling (dir.), Modelling complex ecological dynamics (p. 29-40). Berlin : Springer.
Eveleigh, E. S., McCann, K. S., McCarthy, P. C., Pollock, S. J., Lucarotti, C. J., Morin, B., . . . Faria, L. D. B. (2007). Fluctuations in density of an outbreak species drive diversity cascades in food webs. Proceedings of the National Academy of Sciences of the United States of America, 104(43), 16976-16981. doi : 10.1073/pnas.0704301104
Forrester, J. W. (1961). Industrial dynamics. Waltham : Pegasus Communications.
Forrester, J. W. (1969). Urban dynamics. Waltham : Pegasus Communications.
Forrester, J. W. (1971). World dynamics. Cambridge : Wright-Allen Press.
Jørgensen, S. E. (1986). Fundamentals of ecological modelling. Amsterdam : Elsevier Science Publishers B.V.
Jørgensen, S. E. et Fath, B. D. (2011). Spatial modelling. Dans Fundamentals of ecological modelling (p. 347-368). Oxford : Elsevier.
Malthus, T. R. (1798). An essay on the principle of population. Library of Economics and Liberty. Tiré du web le 15 septembre 2014 : https://www.econlib.org/library/Malthus/malPop.html.
Meadows, D. H., Meadow, D. L., Randers, J. et Behrens III, W. W. (1972). The limits to growth. New York : Signet New American Library.
Odum, E. P. (1953). Fundamentals of ecology. Philadelphia : W.B. Saunders Company.
Patten, B. C. (1971). Systems analysis and simulation in ecology. New York : Academic Press.
Roughgarden, J. (1979). Theory of population genetics and evolutionary ecology: An introduction. New York : Macmillan Publishing Company.
Sholto Maud. (2007). Energy Systems Symbols. Repéré à https://en.wikipedia.org/wiki/File:Energy_Systems_Symbols_H.gif, licence : https://creativecommons.org/licenses/by-sa/3.0/
Thompsma. (2010). A freshwater aquatic and terrestrial food-web. Repéré à https://commons.wikimedia.org/wiki/File:FoodWeb.jpg, licence : https://creativecommons.org/publicdomain/zero/1.0/deed.en
von Bertalanffy, L. (1968). General system theory. New York : George Braziller.